
The barn owl, Tyto alba, is a ubiquitous species that is spread across much of the world, although it prefers more temperate climates. The only continent on which barn owls are not found is Antarctica. Tyto alba is a nocturnal predator that typically hunts small rodents with its heightened senses of sight and hearing. Cliffsides are postulated to be the original habitat for barn owls; however, these birds now are located primarily in rural environments where there is an abundance of prey, along with numerous open buildings in which to nest (Johnsgard, 2002; Marti, 1992).
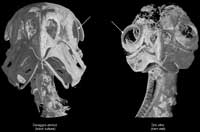
Owls have evolved an astounding sense of both sight and hearing. This is especially true in nocturnal owls such as Tyto alba. Their sight is enhanced by several factors, some of which are directly observable in the skeletal system. Owls possess extremely large eyes, which allow them to gather the maximum amount of light from their environment; therefore, the bony orbits that surround the eyes must be enlarged as well. The orbits of owls are rotated forwards so that they sit anteriorly. This placement differs from the normal condition found in other birds in which the orbits are laterally positioned. Anteriorly facing orbits allow the visual field of each eye to overlap the other, thus, creating an area of binocular vision. The large bony sclerotic rings which sit within the sclera of the eye prevent rotation of the eye itself; accordingly, owls must rotate their entire heads to see to either side of them. In addition to the abovementioned characteristics, owls also possess large corneal surfaces and pupillary areas (in order to gather light) and an extremely high retinal rod to cone ratio (to increase visual sensitivity in dark environments) (Johnsgard, 2002).
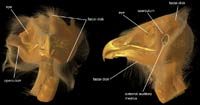
Nocturnal owls also have a remarkable ability to capture prey in the absence of any type of visual signal, using primarily their sense of hearing to guide them. They use soft-tissue structures called facial discs (circular in strigids, and heart-shaped in tytonids) to amplify the sounds traveling to their ears. Fleshy ear flaps (or opercula) also are present. Another unique adaptation of owls is the asymmetry of their external auditory meatus (external ear openings). This asymmetry causes sound to reach each ear at different times (just milliseconds apart), which provides a three-dimensional hearing space. This allows owls to localize their prey with incredible precision (Johnsgard, 2002).
Two groups comprise Strigiformes (owls), the Tytonidae (barn and bay owls) and Strigidae (all other owls) (Randi et al., 1991). Many recent phylogenetic analyses of Aves place the owls as the sister taxon to Accipitridae (hawks, owls, and kites) and Falconidae (caracaras and falcons) (e. g., Mayr et al., 2003; Mayr and Clarke, 2003). This arrangement of evolutionary relationships is supported by a number of diagnostic characters that are observable in the skeleton and musculature of the hindlimb. Other phylogenetic trees, however, show either a polyphyletic Strigiformes or a sister-group relationship between Strigiformes and Caprimulgiformes (see Sibley and Ahlquist, 1990; Bleiweiss et al., 1994; Johnsgard, 1998; Livezey and Zusi, 2001).
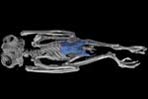
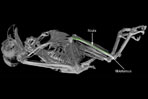
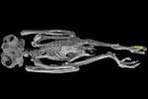
Animations of the burrowing owl, Athene cunicularia, highlighting postcranial osteological characters used to unite owls with Falconidae and Accipitridae (following Mayr et al., 2003 and Mayr and Clarke, 2003):
Pelvis: |
Fibula: |
Tarsometatarsus: |
Phalanges: |
 |
 |
 |
 |

Literature
Baumel, J.J. (ed.). 1993. Handbook of Avian Anatomy : Nomina Anatomica Avium (2nd ed.). Nuttall Ornithological Club, 779 pp.
Beddard, F. E. 1888. On the classification of the Striges. Ibis, series 6, 5:334-344.
Bleiweiss, R., J. A. W. Kirsch, and F.-J. Lapointe. 1994. DNA-DNA hybridization-bsed phylogeny for "higher" nonpasserines: reevaluating a key portion of the avian family tree. Molecular Phylogenetics and Evolution 3:248-255.
Johnsgard, P.A. 2002. North American owls: biology and natural history. Smithsonian Inst. Press, Washington, D. C., 298 pp.
Livezey, B. C., and R. L. Zusi. 2001. Higher-order phylogenetics of modern Aves based on comparative anatomy. Netherlands Journal of Zoology 51:179-205.
Kaup, J. J. 1862. Monograph of the Strigidae. Transactions of the Zoological Society of London 4:201-260.
Marti, C. D. 1992. Tyto alba: barn owl. Birds of North America 8:1-15.
Mayr, G., and J. Clarke. 2003. The deep divergences of neornithine birds: a phylogenetic analysis of morphological characters. Cladistics 19:527-553.
Mayr, G., A. Manegold, and U. S. Johansson. 2002. Monophyletic groups within 'higher land birds' - comparison of morphological and molecular data. Journal of Systematic and Evolutionary Research 41:233-248.
Randi, E., G. Fusco, R. Lorenzini, and F. Spina. 1991. Allozyme divergence and phylogenetic relationships within the Strigiformes. Condor 93:295-301.
Sibley, C. G., and J. E. Ahlquist. 1990. Phylogeny and Classification of Birds: A Study in Molecular Evolution. Yale University Press, New Haven, Connecticut, 976 pp.
Links
Images of Tyto alba on Wildlife Web.
Images and calls of T. alba on The Owl Pages.com.
Information about T. alba on the Animal Diversity Web (Univ. of Michigan Museum of Zoology).
More images and calls of T. alba on Owling.com.



 )


