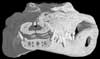
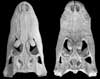
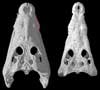
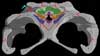
Expert annotations for this species! See below. Expert annotations for this species! See below.
Introduction
|
Crocodylus moreletii is one of four currently recognized species of New World Crocodylus, the genus that includes nearly half of the worlds extant species of crocodylians. This species is commonly known as the Belize crocodile, Morelets crocodile, the Central American crocodile, cocodrilo de pantano, or, confusingly, 'alligator' (Brazaitis, 1973; Ross, 1998). |
Morelets crocodile is relatively small, growing to only 2.5 to 3 meters (approximately 7 to 9 feet) in length (Brazaitis, 1973). Since half of this length is tail, and only about one-sixth is head, these animals are not as intimidating as the larger, longer-snouted American crocodile (Crocodylus acutus), or the more familiar Nile crocodile (C. niloticus) or Indo-Pacific saltwater crocodile (C. porosus).
First described in 1851 (Duméril and Duméril, 1851), Morelets crocodile was not accepted as a distinct species by many workers (Neill, 1971). This was complicated by the suggestion (Barbour and Ramsden, 1919) that the original specimens had actually been collected from Cuba and belonged to the Cuban species Crocodylus rhombifer. Crocodylus moreletii was restored to scientific acceptance by Karl Schmidt (1924), who demonstrated that these animals are indeed found in Belize (then British Honduras), and are distinct from the American crocodile, also found in that country. Despite formal recognition, little was known of the natural history of this animal for most of the twentieth century. Hunting for the leather industry severely depleted many populations to the point that Neill (1971) predicted that C. moreletii would be extinct before any detailed information could be gathered.
Fortunately, this has not occurred; today the species survives, in depleted numbers, in the Caribbean drainages of southern Mexico, Guatemala, and Belize (Ross, 1998; Platt and Thorbjarnson, 2000). Reports of Morelets crocodile from Honduras are disputed (Wilson et al., 1986) and generally disregarded (Ross, 1987). The species is currently protected in all three countries, although hunting continues (Groombridge, 1987). It is listed as 'low risk' but 'conservation dependent' by the IUCN (Ross, 2000), and is listed in CITES Appendix I (Ross, 1998). Efforts to breed the species in captivity have proved successful, and there have been efforts toward farming and other 'sustainable use' harvesting for the leather industry (Ross, 1998).
Natural History and Ecology
Around twenty to forty eggs (Greer, 1975) are laid in a nest formed of vegetable debris piled in a mound (Greer, 1970) during late spring (between April and June, Perez-Higareda, 1980), just before the beginning of the rainy season. Incubation takes approximately 80 days (Britton, 1999). Like most crocodylians, the mother remains in the vicinity of the nest, and will defend it from intruders. She also responds to the chirping of the young in their eggs, and will help them exit the egg, another common crocodylian behavior. Although almost all mother crocodylians will protect their newly-hatched young for at least a short period, Morelets crocodile is one of the few species in which both males and females have been observed participating in defending hatchlings from predators, including larger crocodiles (Rainwater et al., 2000).
The young reach sexual maturity at seven to eight years of age (Platt and Thorbjarnson, 2000). As with many crocodylian species, during this time their diet will change from smaller prey, generally invertebrates and fish, to gradually taking larger prey, including snails, larger fish and mammals, as well as turtles, birds, and other reptiles (Alvarez de Toro, 1974; Britton, 1999). They will take domestic animals, but they have not been reported to be a significant threat to man.
Crocodylus moreletii largely inhabits freshwater environments, including rivers, lakes and marshes, but it will venture into brackish water as well (Ross, 1998; Platt and Thorbjarnson, 2000). The range of Morelets crocodile overlaps that of the C. acutus, with the latter being more common on offshore islands in Belize (Platt and Thorbjarnson, 2000), and in other high-salinity habitats throughout its range. However, like C. acutus, C. moreletti possesses salt-excreting glands on its tongue (Taplin et al., 1985), suggesting that this distribution does not reflect a physiological constraint. In both the Caribbean and the Pacific, there is a ubiquitous, largely brackish-water species that ranges over most of the region (C. acutus and C. porosus, respectively), and a number of geographically restricted, predominantly freshwater forms (Neill, 1971). There is some indication that this pattern is the result of repeated radiations by sea (e.g., Sill, 1968, Taplin et al., 1982; Taplin and Grigg, 1989; Brochu, 2000, 2001a), with a cosmopolitan species occupying a region, then becoming fragmented and isolated and subsequently forming new species, followed by the radiation of another cosmopolitan species that occupies the coastal regions and becomes a barrier to dispersal of the endemic species (ONeill, 1971). In such a scenario, Morelets crocodile would be the older, endemic species being 'bottled up' by the cosmopolitan American crocodile. In Belize, C. moreletiis expansion into brackish-water habitats where one would expect to find C. acutus may be a result of opportunistic expansion following overhunting of both species (Platt and Thorbjarnson, 2000).
Current research on Morelets crocodile is centered on Belize, including research by Steven Platt (Rainwater et al., 2000; Platt and Thorbjarnson, 2000; Ray et al., 2004), Howard Hunt and colleagues (Ross, 1998) and Llewellyn Densmore and colleagues (Rainwater et al., 2000; Dever and Densmore, 2001; Ray et al., 2004). Some of this research has been featured on the National Geographic Channels Reptile Wild program.
Phylogenetic Relationships
The group we call Crocodylia encompasses a lineage of extant species including the true crocodiles, alligators, caimans and gharials. Crocodylians are not lizards; they are archosaurs, a group that also includes the dinosaurs and their living descendants, the birds. Crocodylians are, in fact, the closest living relatives of the birds. There are approximately 23 living species; the uncertainty is due to questions about the distinctiveness of some populations, and the actual number could turn out to be 30 or more (based on the number of subspecies and work currently in progress). Commonly labeled 'living fossils' because of their superficial resemblance to forms that lived alongside the dinosaurs of the Mesozoic Era, crocodylians are actually an evolutionarily dynamic group (Brochu, 2001b, 2003). The small number of living species, and their superficial resemblance to ancient crocs, is almost certainly a result of their historically limited ecological role; there are only so many places a large-bodied, amphibious, largely fresh-water predator can live, and each place can only hold so many individuals. Fossil relatives include fully aquatic predators, terrestrial predators, and even terrestrial herbivores, and croc diversity was correspondingly higher in the past.
Morelets crocodile belongs to the genus Crocodylus, the true crocodiles. Species of Crocodylus, except the African slender-snouted crocodile (C. cataphractus), can be recognized by their distinctive cluster of six enlarged nuchal scales, four scales across in front and two behind, over the neck. These are separated from the scales of the back by a patch of very fine scales between the forelimbs, sometimes with one or two pairs of relatively smaller plate-like scales as well.
There appear to be at least two natural groups within Crocodylus, an American group and an Indo-Pacific group (Brochu, 2000); the former includes C. moreletii. The relationship of C. niloticus to these two groups is uncertain, although it shows superficial similarities to the New World species. Crocodylus cataphractus lies outside the other species.
The New World crocodiles are characterized by an elevation of the nasal bones, a pair of bones that lie along the midline of the snout. This can give the head a distinctive 'flat-topped' or slightly humped profile. In some animals (but not TMM M-4980), this elevation is taken to an extreme, forming a distinct bulge in front of the eyes. This can give the animal a 'roman nose' profile, which is most pronounced in C. acutus.
There is DNA evidence of interbreeding between Crocodylus moreletii and C. acutus in Belize (Ray et al., 2004). While this may seem at odds with some ideas about what a species is, it is not at all uncommon in nature. In fact, hybridization also occurs between C. rhombifer and C. acutus, at least in captivity. Under singular, quite artificial circumstances, fertile hybrids have even been produced by C. rhombifer and the Siamese crocodile, C. siamensis, species that live on opposite sides of the world (Ross, 1998). Among some species of organisms, occasional hybridization is thought to strengthen a population by introducing new genes (more properly, new alleles), a process called introgression. It is not presently clear to what extent introgression or outright hybridization may have affected crocodylian evolution.
Fossil Record
There are two reports of fossil Crocodylus moreletii. The first, from the mid(?)-Pleistocene of Guatemala, is based on a maxilla (upper jaw), and was used as the basis for a new subspecies, C. m. barnumbrowni (Mook, 1959). Mook (1959) saw similarities between his specimen and C. moreletti, including a strong angulation of the side of the upper jaw around the tenth tooth from the tip of the nose (subsequently emphasized by Neill, 1971). The shape of the premaxillary-maxillary suture on the palate is also like that of C. moreletii. There has been no further review of this specimen. There is no way to tell whether or not this specimen truly was a member of the Recent species. The anatomical evidence suggests a close relationship with the modern form, but the specimen is so fragmentary as to make a definitive statement impossible.
A second specimen, a pair of partial lower jaws from Baja California, was referred to Crocodylus cf. C. moreletii by Miller (1980). The 'cf.' is used by some authors to indicate that a specimen is closely related to, but not the same as, the indicated species; others use it to say that it could be that species, but not enough information is available. In this case, the specimen is more similar to Morelets crocodile than it is to other living species, but is different from that species in detail (Miller, 1980). The locality is well outside the range of C. moreletii, which is not present along the Pacific coast; C. acutus does live on the Pacific coast, ranging as far north as the mainland opposite the tip of Baja California. Without further study (or, better still, more specimens) it is not possible to say whether these remains represent Morelets crocodile or a previously unknown, extinct species.
Features Visible in the CT Data
Click on the topics below for more information:
Acknowledgments
I am very grateful to the following individuals and institutions for providing access to specimens or other important materials: Colin McCarthy at the Natural History Museum, London; Georges Lenglet at the Institut Royal des Sciences Naturelles de Belgique, Brussels; Frank Glaw and Dieter Fuchs at the Zoologische Staatssammlung München in Munich; Gunther Koehler and Monika Laudahn at the Senckenberg Museum in Frankfurt; Harold Voris, Alan Resetar, Jamie Ladonski, and Jennifer Mui at the Field Museum of Natural History in Chicago; Chris Brochu at the University of Iowa; Albert Prieto-Marquez at Florida State University; Chris Bell at the University of Texas at Austin; Amy Balanoff and Matt Colbert at The University of Texas High-Resolution X-ray CT Facility; and Tim Rowe, Wann Langston, Pamela Owen, and Lyn Murray at the Texas Memorial Museum in Austin. I thank Jonathan Franzosa for initiating the scanning of this specimen, and for granting me access to the data in advance of its public release. I would also like to thank the Department of Geological Sciences at the University of Texas at Austin, the Center for Computational Biology at UT Austin, and the Texas Memorial Museum for financial support. Funding to visit the collections listed above was provided by the Dorothy Ogden Carsey Memorial Fund, the Johan A. Udden Fund, and the Ronald K. DeFord Field Scholarship Fund of the Geology Foundation at UT Austin, and the J. Nalle Regents Professorship (to Tim Rowe) of the Department of Geological Sciences at UT Austin. This work would not be possible without the diligence and perseverance of the Interlibrary Services Division of the University of Texas Libraries.

About the Species
About this Specimen
This specimen was scanned by Matthew Colbert on 21 August 2003 along the coronal axis for a total of 663 slices. Each slice is 0.5 mm thick, with an interslice spacing of 0.5 mm and a field of reconstruction of 195.0 mm.
About the
Scan
Literature
Alvarez de Toro, M. 1974. Los Crocodylia de Mexico (Estudio Comparativo). Instituto Mexicano de Recursos Naturales Renovables, Mexico D. F. 70 pp.
Barbour, T., and C. T. Ramsden. 1919. The herpetology of Cuba. Memoirs of the Museum of Comparative Zoology 47:71-213.
Bertau, M. 1935. Zur entwicklungsgeschichte des geruchsorgans der krokodile. Zeitschrift für Anatomie und Entwicklungsgeschichte 104:168-202.
Brazaitis, P. 1973. The identification of living crocodilians. Zoologica 58:59101.
Britton, A. 1999. Crocodilians: natural history and conservation [Online]. [2 August, 1999].
Brochu, C. A. 2000. Phylogenetic relationships and divergence timing of Crocodylus based on morphology and the fossil record. Copeia 2000: 657-673.
Brochu, C. A. 2001a. Congruence between physiology, phylognetics and the fossil record on crocodylian historical biogeography, pp. 9-28. In G. C. Grigg, F. Seebacher and C. E. Franklin (eds.), Crocodilian Biology and Evolution, Surrey, Beatty and Sons, Chipping Norton, New South Wales.
Brochu, C. A. 2001b. Crocodylian snouts in space and time: phylogenetic approaches toward adaptive radiation. American Zoologist 41:564-585.
Brochu, C. A. 2003. Phylogenetic approaches toward crocodylian history. Annual Review of Earth and Planetary Sciences 31:357-397.
Colbert, E. H. 1946. The eustachian tubes in the Crocodilia. Copeia 1946:11-14.
Deraniyagala, P. E. P. 1939. The Tetrapod Reptiles of Ceylon, Volume I. Testudinates and Crocodilians. Dulau and Company, London. 412 pp.
Dever, J. A., and L. D. Densmore, III. 2001. Microsatellites in Morelets crocodile (Crocodylus moreletii) and their utility in addressing crocodilian population genetics questions. Journal of Herpetology 35:541-544.
Duméril, A. M. C., and A. H. A. Duméril. 1851. Catalogue méthodique de la collection des reptiles du Muséum d'Histoire Naturelle de Paris. Gide and Baudry, Paris. iv + 224 pp.
Edmund, A. G. 1962. Sequence and rate of tooth replacement in the Crocodilia. Contributions of the Royal Ontario Museum, Life Sciences 56:7-42.
Greer, A. E. 1970. Evolutionary and systematic significance of crocodilian nesting habits. Nature 227:523-524.
Greer, A. 1975. Clutch size in crocodilians. Journal of Herpetology 9:319-322.
Groombridge, B. 1987. The distribution and status of world crocodilians, pp. 9-21. In G. J. W. Webb, S. C. Manolis, and P. J. Whitehead (eds.), Wildlife Management: Crocodiles and Alligators, Surrey, Beatty and Sons, Chipping Norton, New South Wales.
Hall, P. M. 1989. Variation in geographic isolates of the New Guinea crocodile (Crocodylus novaeguineae Schmidt) compared with the similar, allopatric, Philippine crocodile (Crocodylus mindorensis Schmidt). Copeia 1989:7180.
Hall, P. M., and K. M. Portier. 1994. Cranial morphometry of New Guinea (Crocodylus novaeguineae) crocodiles: ontogenetic variation in relative growth of the skull and an assessment of its utility as a predictor of the sex and size of individuals. Herpetological Monographs 8:203225.
Iordansky, N. N. 1973. The skull of the Crocodilia, pp. 201-260. In C. Gans and T. Parson (eds.), Biology of the Reptilia, vol. 4, Academic Press, London.
Kälin, J. A. 1933. Beiträge zur vergleichenden Osteologie des Crocodilidenschädels. Zoologisches Jahrbucher 57:535-714.
Kälin, J. A. 1937. Über skeletanomalien bei crocodiliden. Zeitschrift für Morphologie und Ökologie der Tiere 32: 327-347.
Meers, M. B. 1996. Three-dimensional analysis of differences in cranial morphology between captive and wild American alligators [Online]. [accessed September 28, 2005].
Miller, W. E. 1980. The late Pliocene Las Tunas Local Fauna from southernmost Baja California, Mexico. Journal of Paleontology 54:762-805.
Mook, C. C. 1921. Individual and age variation in the skulls of Recent Crocodilia. Bulletin of the American Museum of Naural History 44:51-66.
Mook, C. C. 1959. A new Pleistocene crocodilian from Guatamala. American Museum Novitates 1975:1-6.
Neill, W. T. 1971. The Last of the Ruling Reptiles: Alligators, Crocodiles, and their Kin. Columbia University Press: New York. 486 pp.
Owen, R. 1850. On the communications between the cavity of the tympanum and the palate in the Crocodilia (gavials, alligator and crocodiles). Philosophical Transactions of the Royal Society of London 27:521-527.
Perez-Higareda, G. 1980. Notes on nesting of Crocodylus moreletii in Southern Veracruz, Mexico. Bulletin of the Maryland Herpetological Society 16:52-53.
Platt, S. G., and J. B. Thorbjarnason. 2000. Population status and conservation of Morelet's crocodile. Biological Conservation 96:21-29.
Ray, D. A., J. A. Dever, S. G. Platt, T. R. Rainwater, A. G. Finger, S. T. McMurry, M. A. Batzer, B. Barr, P. J. Stafford, J. McKnight, and L. D. Densmore. 2004. Low levels of nucleotide diversity in Crocodylus moreletii and evidence for hybridization with C. acutus. Conservation Genetics 5:449-462.
Rainwater, T., S. T. McMurry, and S. G. Platt. 2000. Crocodylus moreletii (Morelets crocodile). Behavior. Herpetological Review 31:237.
Ross, C. A. 1987. Crocodylus moreletii. Catalogue of American Amphibians and Reptiles 407:1-3.
Ross, C. A., and F. D. Ross. 1974. Caudal scalation of Central American Crocodylus. Proceedings of the Biological Society of Washington 100:713-716.
Ross, C. A., and F. D. Ross. 1987. Identity of Crocodylus mexicanus Bocourt, 1869 (Reptilia: Crocodylidae). Proceedings of the Biological Society of Washington 100:713-716.
Ross, J. P. (ed.). 1998. Crocodiles. Status Survey and Conservation Action Plan [Online]. 2nd Edition. IUCN/SSC Crocodile Specialist Group. IUCN, Gland, Switzerland and Cambridge, UK. viii + 167 pp. [6 July 1998].
Ross, J. P. 2000. Crocodylus moreletii. In: IUCN 2004. 2004 IUCN Red List of Threatened Species.
Schmidt, K. P. 1924. Notes on Central American crocodiles. Field Museum of Natural History, Zoological Series 12:79-92.
Schute, C. C. D., and A. dA. Bellairs. 1955. The external ear in Crocodilia. Proceedings of the Zoological Society of London 124:741-749.
Sill, W. D. 1968. The zoogeography of the Crocodilia. Copeia 1968:76-88.
Taplin, L. E., and G. C. Grigg. 1989. Historical zoogeography of the eusuchian crocodilians: a physiological perspective. American Zoologist 29:885-901.
Taplin, L. E., G. C. Grigg, and L. Beard. 1985. Salt gland function in fresh water crocodiles: evidence for a marine phase in eusuchian evolution?, pp. 403-410. In G. Grigg, R. Shine, and H. Ehmann (eds.), Biology of Australasian Frogs and Reptiles, Royal Zoological Society of New South Wales.
Taplin, L. E., G. C. Grigg, P. Harlow, T. M. Ellis, and W. A. Dunson. 1982. Lingual salt glands in Crocodylus acutus and C. johnstoni, and their absence from Alligator mississippiensis and Caiman Crocodilus. Journal of Comparative Physiology 149:43-47.
Wegner, R. N. 1957. Die Nebenhölen der nase bei den Krokodilen. Wissenschaftliche Zeitschrift der Ernst Mortiz Arnd-Universität 7:1-37.
Wilson, L. D., J. R. McCranie, and K. L. Williams. 1986. The identity of the crocodile of Lago de Yojoa, Honduras. Journal of Herpetology 20:87-88.
Witmer, L. M. 1995. Homology of facial structures in extant archosaurs (birds and crocodilians), with special reference to paranasal pneumaticity and nasal conchae. Journal of Morphology 225:269-327.
Links
Natural history and images of Crocodylus moreletii on the Crocodilian Species List (Florida Museum of Natural History).
Images of Morelet's crocodile on the Animal Diversity Web (University of Michigan Museum of Zoology).
Literature
& Links
Additional Imagery
 |



 )