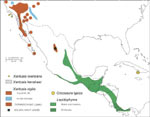
Xantusiidae is a clade of viviparous (live bearing) lizards that ranges from southwestern North America and Baja California (Xantusia) into Central America (Lepidophyma) and Cuba (Cricosaura). Xantusia bezyi is found in rock-crevice habitats in central Arizona. Xantusiidae is a relatively small clade, with 3 genera and approximately 30 living species. Lepidophyma is the most speciose (~18 species), whereas Cricosaura is monotypic. Xantusiids have a reasonably good fossil record extending from the mid-Paleocene onward in western North America. |
|
Xantusiids are fascinating lizards for several reasons. First, although they are almost uniformly diminuitive (Xantusia bezyi measures less than 4 cm snout-vent length, and the largest xantusiid species measure about 10 cm snout-vent length), xantusiids generally take several years to reach sexual maturity, and several species give birth to just 1 or 2 offspring. It is a more usual reproductive strategy for small lizards to mature quickly and produce large numbers of offspring, to increase their chances of survival. Despite this low reproductive potential, xantusiid neonates actually have a high life expectancy; this can be attributed at least in part to their secretive lifestyle, which leads to the second reason why xantusiids are particularly interesting -- microhabitat specialization.
Microhabitat specialization is an ecological hallmark of Xantusiidae. Many populations are narrowly restricted to specific niches -- crevices (e.g., Xantusia henshawi in exfoliating granitic cap rocks), interstices in agaves and yuccas in dry climates (Xantusia vigilis), decaying logs in wet climates (e.g., Lepidophyma flavimaculatum) -- and individuals may be found under the same patch of cover throughout their lives!
These microhabitat restrictions result in extremely disjunct geographical distributions, and also may be responsible for some morphological convergence within the group (e.g., flattened skulls for crevice dwelling). Xantusiidae also includes two insular endemics: the Cuban Cricosaura typica is the only xantusiid found in the West Indies and is interpreted as one of the Caribbean's few ancient endemic vertebrate lineages; and Xantusia riversiana (formerly Klauberina riversiana) is limited to three of the Channel Islands off the coast of California.
The phylogenetic relationships of Xantusiidae are problematic. Morphology and molecules produce different topologies within the clade: morphology recovers a Cricosaura + Lepidophyma clade, while mitochondrial genes recover a Lepidophyma + Xantusia clade. Lack of resolution of relationships within Xantusiidae has hindered the placement of this clade within the squamate tree. Xantusiidae is a "tree-changing" taxon: it causes homoplasy wherever it is placed, and its placement can tip the balance between the two primary competing hypotheses of scleroglossan relationships. Xantusiidae is traditionally placed within Scincomorpha, but some analyses have placed it near Gekkota. Thus, Xantusiidae is either a highly derived, or extremely basal, scleroglossan clade. Previous analyses of squamate phylogeny have almost certainly suffered in relying on species of the readily available -- but relatively derived -- genus Xantusia as exemplars for Xantusiidae. Cricosaura or a species of Lepidophyma would be more appropriate, but both are exceedingly rare in collections; indeed, some species of Lepidophyma are known from only 1 or 2 specimens.
Whatever the placement of Xantusiidae within squamates, there is no doubt that xantusiids are monophyletic. The following are some of the hypothesized synapomorphies of the lineage (from Estes et al., 1988), most of which can be seen in the skull reconstructions above: supratemporal fenestra closed primarily by postorbital; parietals paired well into postembryonic ontogeny; parietal table extensive posteriorly, largely obscuring braincase in dorsal view, supratemporal process short; vomers fused; ectopterygoid contacts palatine anterolaterally, excluding maxilla from suborbital fenestra; ectopterygoid enlarged medially, restricting suborbital fenestra.

About the Species
This specimen was collected in Maricopa County, Arizona. It was made available to the University of Texas High-Resolution X-ray CT Facility for scanning by Dr. Jessie Maisano of The University of Texas and Dr. Jacques Gauthier of Yale University. Funding for scanning was provided by an NSF grant (DEB-0132227) to Dr. Jack Sites of Brigham Young University. Funding for image processing was provided by a National Science Foundation Digital Libraries Initiative grant to Dr. Timothy Rowe of The University of Texas at Austin.
About this Specimen
The specimen was scanned by Matthew Colbert on 15 May 2005 along the coronal axis for a total of 525 1024x1024 pixel slices. Each slice is 0.0252 mm thick, with an interslice spacing of 0.0252 mm and a field of reconstruction of 12 mm.
About the
Scan
Literature
Bezy, R. L. 1982. Xantusia vigilis. Catalogue of American Amphibians and Reptiles 302.1-302.4.
Bezy, R. L. 1988. The natural history of the night lizards, family Xantusiidae, p. 1-12. In H. F. DeLisle et al. (eds.), Proceedings of the Conference on California Herpetology. Southwest Herpetological Society Special Publication 4.
Bezy, R. L. 1989. Night lizards: the evolution of habitat specialists. Terra 28:29-34.
Bezy, R. L., and J. L. Camarillo. 2002. Systematics of xantusiid lizards of the genus Lepidophyma. Los Angeles County Museum Contributions in Science 493:1-41.
Crother, B. I., M. M. Miyamoto, and W. F. Presch. 1986. Phylogeny and biogeography of the lizard family Xantusiidae. Systematic Zoology 35:37-45.
Estes, R. 1983. Sauria Terrestria, Amphisbaenia. Handbuch der Palaoherpetologie, Part 10A. Gustav Fischer Verlag, Stuttgart.
Estes, R., K. de Queiroz, and J. Gauthier. 1988. Phylogenetic relationships within Squamata, p. 119-281. In R. G. Estes and G. K. Pregill (eds.), Phylogenetic Relationships of the Lizard Families: Essays Commemorating Charles L. Camp. Stanford University Press, Stanford.
Fellers, G. M., and C. A. Drost. 1991. Ecology of the island night lizard, Xantusia riversiana, on Santa Barbara Island, California. Herpetological Monographs 5:28-78.
Hedges, S. B., R. L. Bezy, and L. B. Maxson. 1991. Phylogenetic relationships and biogeography of xantusiid lizards, inferred from mitochondrial DNA sequences. Molecular Biology and Evolution 8:767-780.
Lee, M. S. Y. 1998. Convergent evolution and character correlation in burrowing reptiles: towards a resolution of squamte relationships. Biological Journal of the Linnean Society 63:369-453.
Macey, J. R., A. Larson, N. B. Ananjeva, and T. J. Papenfuss. 1997. Evolutionary shifts in three major structural features of the mitochondrial genome among iguanian lizards. Journal of Molecular Evolution 44:660-674.
Savage, J. M. 1955. The lizard family Xantusiidae: an evolutionary study. Ph.D. Dissertation, Stanford University.
Savage, J. M. 1963. Studies on the lizard family Xantusiidae. IV. The genera. Los Angeles County Museum Contributions in Science 71:3-38.
Sinclair, E. A., Bezy, R. L., Bolles, K., Camarillo R., J. L., Crandall, K. A. and J. W. Sites Jr. 2004. Testing species boundaries in an ancient species complex with deep phylogeographic history: Genus Xantusia (Squamata: Xantusiidae). The American Naturalist 164:396-414.
Van Denburgh, J. 1895. The species of the genus Xantusia. Proceedings of the California Academy of Sciences (Series 2) 5:523-534.
Zweifel, R. G., and C. H. Lowe. 1966. The ecology of a population of Xantusia vigilis, the desert night lizard. American Museum Novitates 2247:1-57.
Links
Xantusiidae page on the EMBL Reptile Database
pictures of Xantusia bezyi from the Tucson Herpetological Society
Literature
& Links
Three-dimensional volumetric renderings of the skull with the scleral ossicles, hyoid and jaw removed, and of the isolated left mandible. All are less than 2mb.
Additional Imagery
 |



 )